Introduction
The most common endocrine malignancy and the fifth most common cancer type worldwide is thyroid cancer, which is increasing (>5% per year).
1 Thyroid carcinoma has several factors contributing to its onset and progression, although the exact etiology is still under investigation. These factors include ionizing radiation exposure, environmental pollution, smoking, hormonal imbalances, metabolic diseases such as obesity, and gender.
2,3 Histopathology has identified several subtypes of thyroid carcinoma, with papillary thyroid carcinoma (PTC) accounting for 85% to 90% of all thyroid carcinomas.
4,5 Other thyroid cancer subtypes include follicular thyroid carcinoma, anaplastic thyroid carcinoma, and medullary thyroid carcinoma.
6 Despite a favorable long-term prognosis, the relatively high recurrence rate of PTC presents a significant challenge, especially when it comes to diagnosing and treating metastatic recurrence in advanced cases.
7,8 In addition, overdiagnosis and overtreatment are common, especially in indolent cases. Identifying and treating indolent thyroid cancer cases can expose patients to unnecessary risks.
9 Molecular-based studies can aid in diagnosing and treating PTC by providing insight into its underlying mechanisms.
10,11 Consequently, there is an urgent and compelling need to develop novel and highly sensitive biomarkers that accurately identify specific PTC patients. This will enable personalized treatment strategies.
Lysophosphatidic acid receptor 1 (LPAR1) is a member of the G protein-coupled receptor family that binds to lysophosphatidic acid (LPA), a small phospholipid commonly found in serum, ascitic effusions, and inflammatory fluids.
12 LPAR1 is pivotal to numerous biological functions, including chemotaxis, cell proliferation, differentiation, platelet aggregation, and tumor progression.
13-16 Multiple cancer cell lines and primary tumors exhibit abnormal LPAR1 expression.
17 LPAR1 has been identified as a prognostic biomarker for different types of cancer in several studies. They emphasize its crucial involvement in prostate, gastric, and pancreatic cancers.
12,18-20 However, the specific functions and mechanisms of LPAR1 in tumor hallmarks, such as epithelial-mesenchymal transition (EMT), remain largely unknown. EMT, a process in which epithelial cells acquire mesenchymal characteristics, occurs in various biological processes. EMT plays a significant role in promoting tumor malignancy by enhancing invasiveness and metastatic potential.
21However, the biological role of LPAR1 in PTC remains unknown. In this study, we sought to investigate the expression of LPAR1 in PTC patients undergoing EMT by analyzing data from The Cancer Genome Atlas (TCGA) and the National Center for Biotechnology Information Gene Expression Omnibus (GEO) databases. Through comprehensive multidimensional analysis, we explored the genomic functional networks associated with LPAR1 in PTC and delved into its potential role in EMT. Our results shed light on LPAR1 as a promising and novel target for diagnosing and treating PTC. This highlights its significance in furthering our understanding of the disease.
Materia and Methods
RNA sequencing data preprocessing
On August 12, 2023, we accessed the GDC data portal (
https://portal.gdc.cancer.gov/) to obtain RNA-Seq data and clinical information related to the TCGA thyroid carcinoma project (project ID = TCGA-THCA). The dataset consisted of 505 primary tumors and 59 adjacent non-tumor tissues for mRNA analysis. We removed duplicate samples and those without complete clinical-TNM stage data, resulting in a final count of 279 samples (203 females and 76 males) for primary tumors and 59 samples (41 females and 17 males) for adjacent non-tumor tissues.
Expression analysis of RNAseq data related to the THCA project
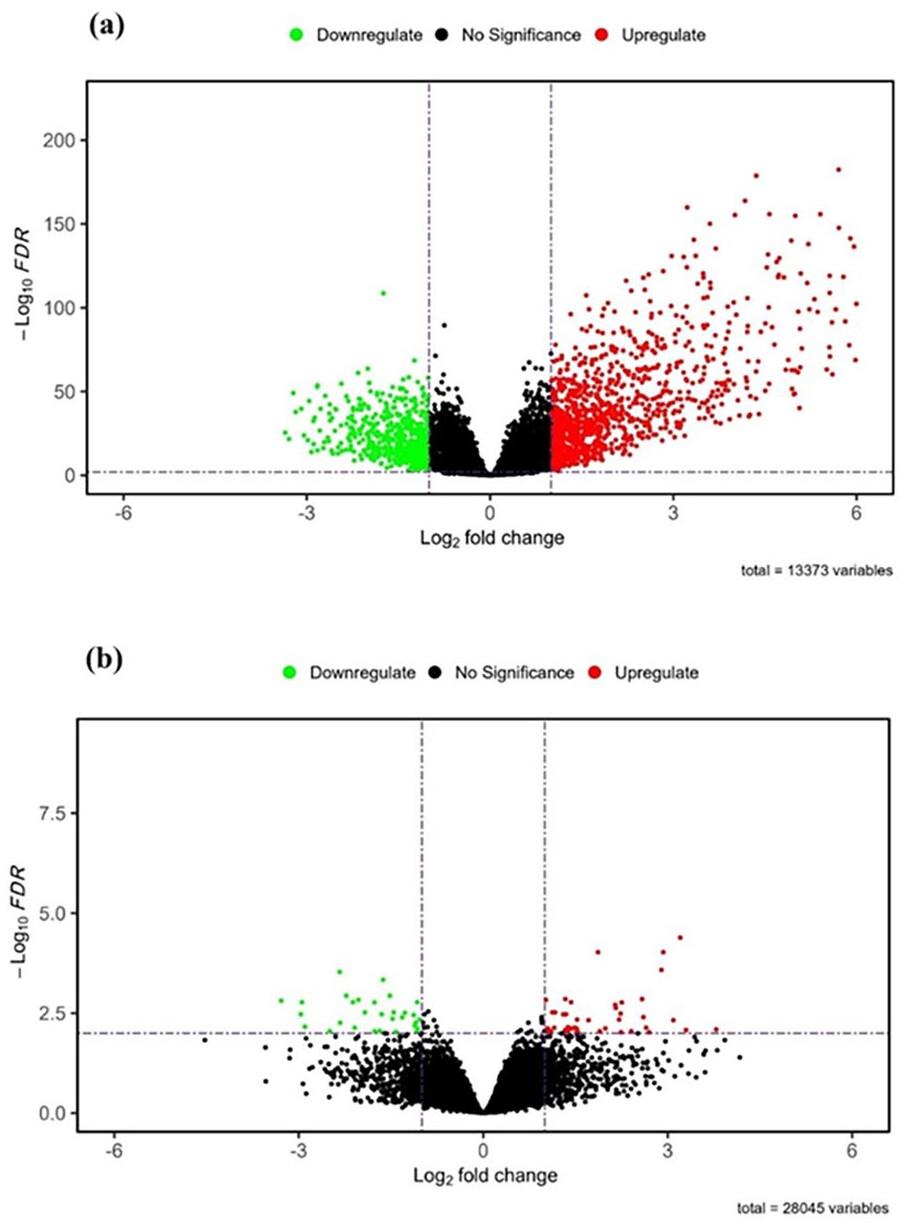
We used the DESeq2 package in R to conduct mRNA expression analysis, specifically comparing PTC samples with normal ones. Our analysis assessed differential expression between primary tumor and normal tissue samples. Genes were considered differentially expressed if they satisfied the criteria of a False Discovery Rate (FDR) of less than 0.01 and an absolute log2 Fold Change (logFC) greater than 1.
Identification of proteins related to epithelial-to-mesenchymal transition (EMT)
To investigate the role of proteins in EMT, we analyzed the GSE6004 dataset obtained from the Gene Expression Omnibus (GEO). The analysis was performed using the GEOquery package in R. This dataset comprised gene expression of EMT in PTC invasion, including 18 samples (four normal tissues and 14 thyroid cancer tissues). We employed the limma method to identify differentially expressed genes associated with EMT. We considered significance based on an adjusted p-value (Adjpval) threshold of less than 0.01 and an absolute log fold change (logFC) greater than 1.
Selection of LPAR1 as the main study target
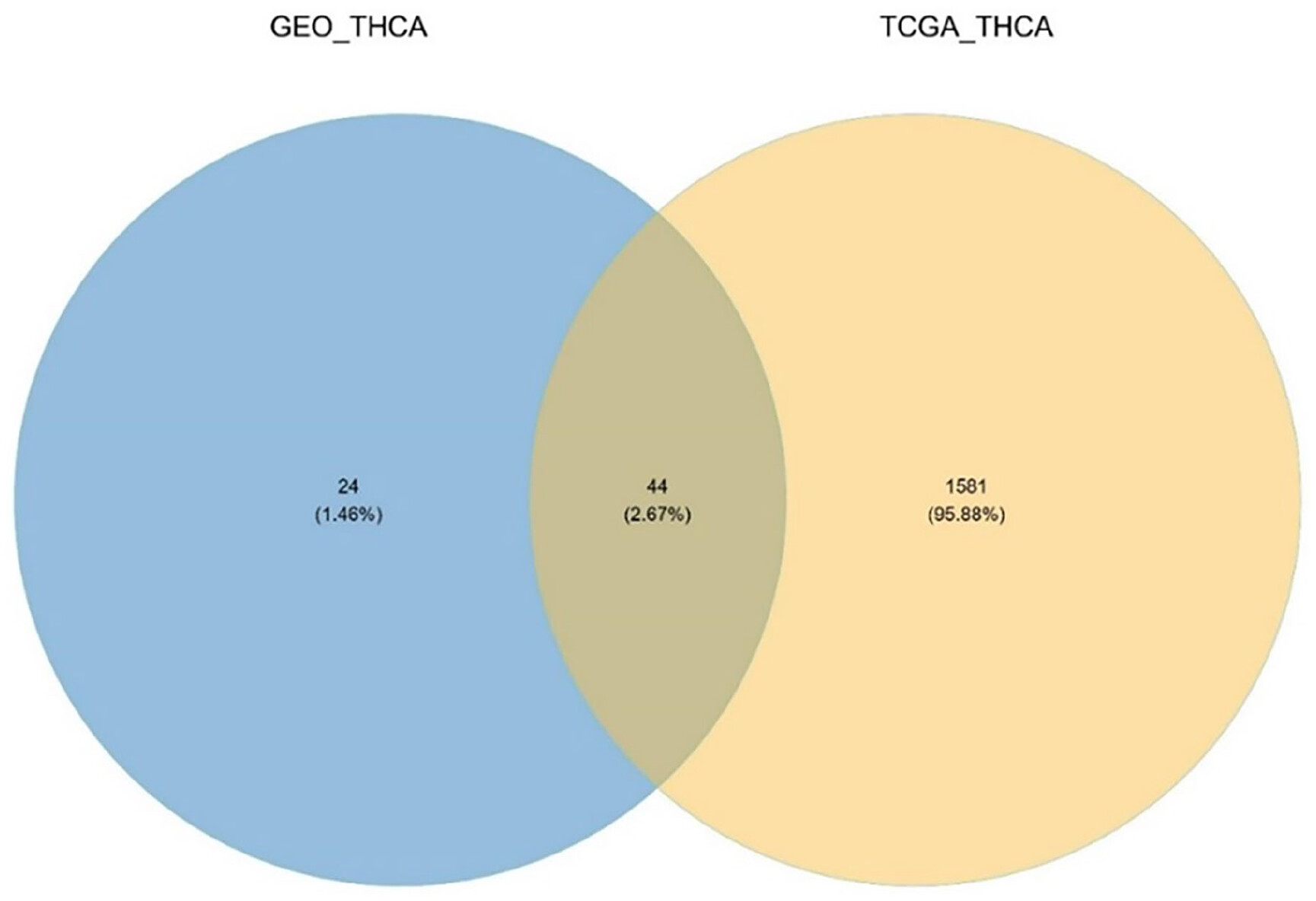
We adopted a comprehensive approach by intersecting the results of RNA expression analysis obtained from the TCGA-THCA dataset with those derived from the GEO dataset. Through this analysis, we identified LPAR1 as a target of interest for further investigation and analysis.
Protein-protein interaction (PPI) analysis
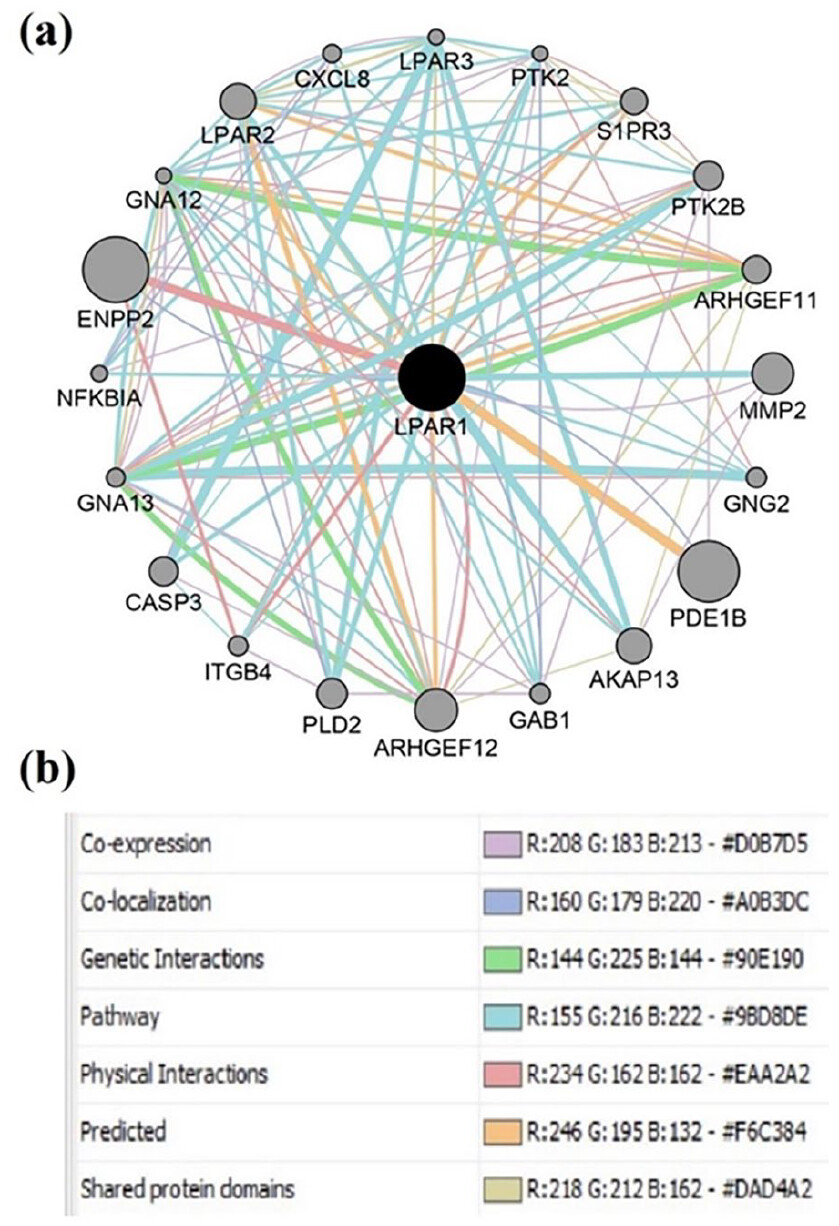
To better understand LPAR1 protein-protein interactions and their potential to relate to EMT, we utilized the GeneMANIA database. GeneMANIA serves as a valuable resource for exploring functional associations between genes. Through this powerful database, we explored the protein-protein interactions between LPAR1 and its relevant partners. The GeneMANIA database integrates diverse sources of applicable association data, including genetic interactions, co-expression patterns, pathways, and physical interactions. We aimed to uncover a more comprehensive view of the interactions within the protein-protein interaction (PPI) network involving LPAR1 by harnessing GeneMANIA database capabilities.
Functional enrichment analysis of PPI network
To gain a deeper understanding of the functions and enriched pathways associated with the proteins identified in the protein-protein interaction (PPI) network, we performed gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses. The GO analysis encompasses 3 aspects: cellular component (CC), biological process (BP), and molecular function (MF). To conduct the GO analysis, we utilized the ClusterProfiler R package (Version 4.0.2) and employed the Benjamini-Hochberg method with a significance threshold of P-value < .05 and q-value < .05. This allowed us to identify the functional categories and molecular functions associated with the proteins. Additionally, the KEGG analysis revealed the potential roles of these genes within specific pathways.
Immune infiltration analysis
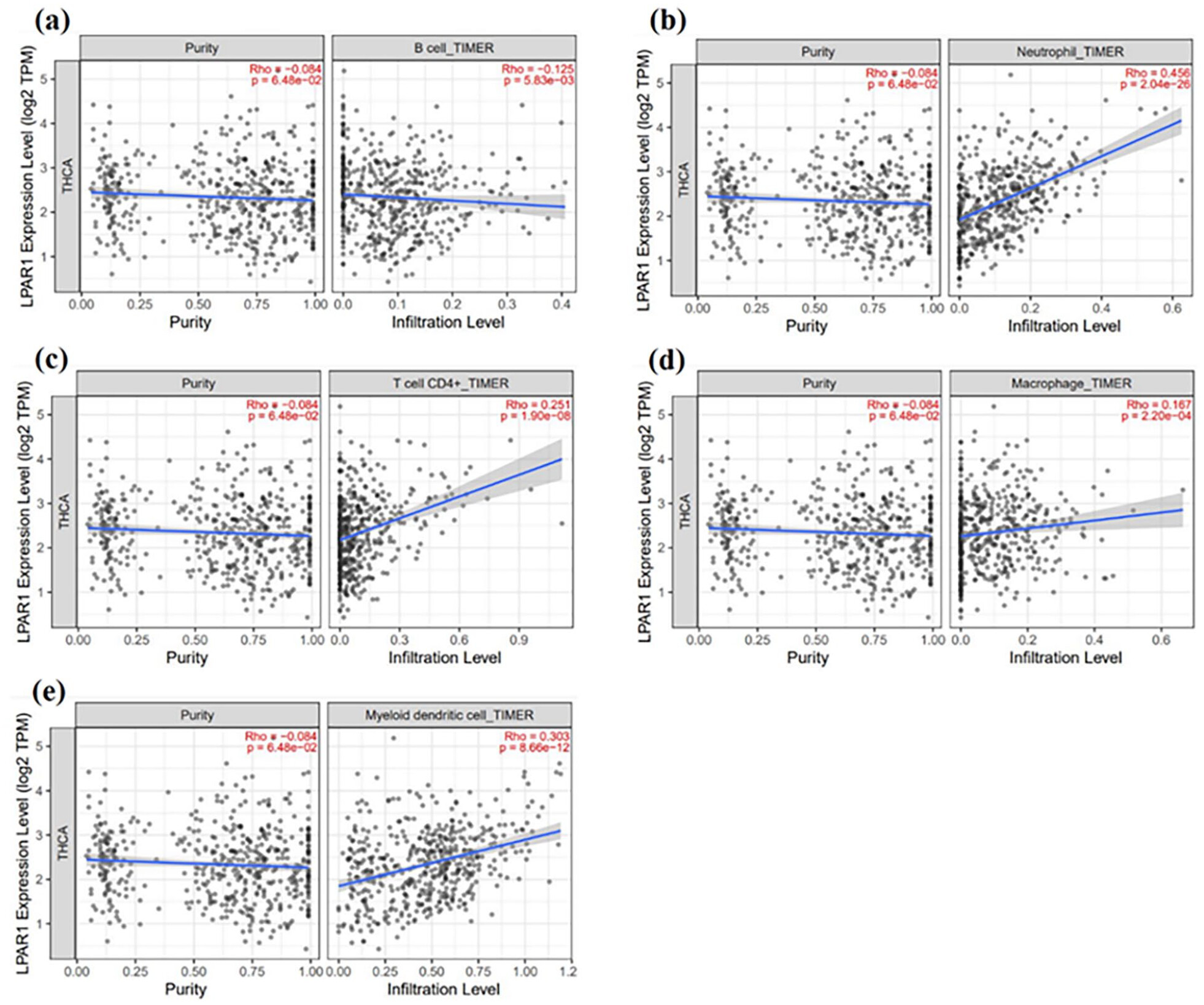
To investigate the potential correlation between LPAR1 expression and immune infiltrates in PTC, we utilized the Tumor Immune Estimation Resource (TIMER). This online tool leverages TCGA data to comprehensively analyze immune infiltrates across various cancer types. We computed the abundance of immune infiltrates, including CD8+ T cells, CD4+ T cells, dendritic cells, neutrophils, B cells, and macrophages, within PTC samples using partial deconvolution and linear least square regression. We aimed to establish a connection between LPAR1 expression levels and immune infiltrates in PTC samples. This was done by exploring potential associations between LPAR1 and the immune microenvironment in PTC.
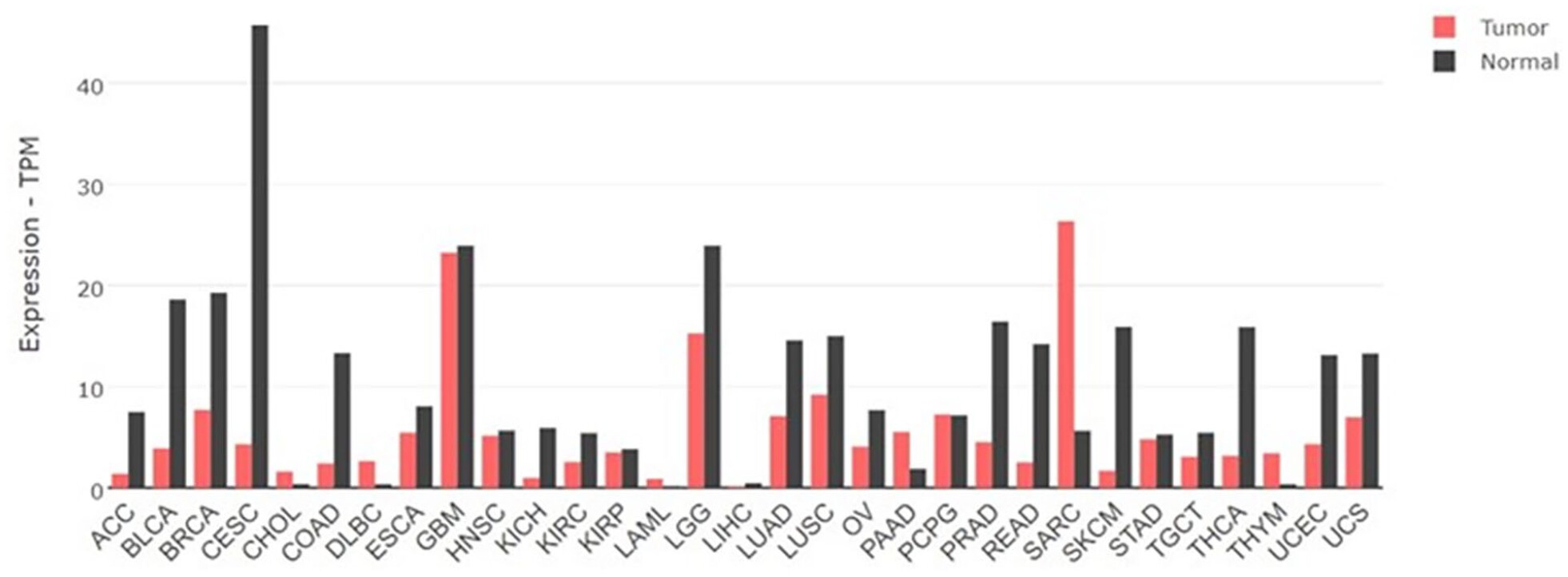
Evaluation of LPAR1 expression in various cancers through the Gepia2 database
To expand our knowledge of LPAR1 expression across various types of cancer, we utilized the Gepia2 database (
http://gepia2.cancer-pku.cn/). Gepia2 is a valuable tool for large-scale RNA sequencing and expression analysis. This database integrates data from TCGA and the Genotype-Tissue Expression (GTEx) projects. By accessing the Gepia2 database, we explored and examined LPAR1 expression levels in various cancer types.
Prediction of miRNAs related to LPAR1
Our study utilized the multiMiR tool in the R programing language to predict miRNA target genes and investigate miRNA-mRNA interactions. The multiMiR tool, available through its web server at
http://multimir.org/, consolidates a wide range of predicted and validated miRNA-target interactions. It incorporates numerous databases, providing a comprehensive selection of interactions based on various binding strength thresholds, including those associated with diseases and medications. Our research focused on the TarBase database within multiMiR, which offers verified miRNA-target interactions.
Cell culture
The Nthy-ori 3-1 human thyroid epithelial cell line was obtained from the American Type Culture Collection (Manassas, VA, USA), while the BCPAP human papillary thyroid carcinoma cell line was obtained from the Chinese Academy of Sciences. Both cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen Technologies, USA) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 µg/mL streptomycin. The cells were incubated at 37°C in a humidified atmosphere with 5% CO2.
RNA isolation, cDNA synthesis, and RT-qPCR
Cell lines were subjected to RNA isolation using TRIzol (Life Technologies, USA) following the manufacturer’s instructions. Subsequently, RNA was reverse transcribed into cDNA using the PrimeScript RT kit (Takara Biotechnology). The expression of LPAR1 was assessed by RT-qPCR on a Light Cycler 480II real-time PCR system (Roche Diagnostics, Switzerland) using SYBR Premix Ex Taq™ (Takara, Dalian, China) as the detection method, with GAPDH serving as the endogenous control. The 2−∆∆Cq method was utilized to determine the relative expression values, enabling analysis of the gene expression changes observed in the real-time quantitative PCR experiments. The primer sequences employed in our study are provided below: LPAR1 (F: 5’- CTCATTGACACCACCGTGAC-3’; R: 5’-GTTGAAAATGGCCCAGAAGA-3’), GAPDH (F: 5’-GTCTCCTCTGACTTCAACAGCG -3’; R: 5’-ACCACCCTGTTGCTGTAGCCAA-3’).
Statistical analysis
The data were analyzed using GraphPad Prism 5.0 statistical software (GraphPad Software, Inc., La Jolla, CA, USA). Statistical differences between groups were evaluated using an unpaired t-test. A significance level of P < .05 was used to determine statistical significance. To ensure robustness and reliability, all experiments were performed in duplicate with at least 3 replications, which minimized the chance of chance variations, thus strengthening the validity of the findings.
Discussion
The onset of EMT in thyroid epithelial cells results in the aberrant regulation of multiple factors, such as transcription factors, signaling cascades, epigenetic changes, and the tumor environment. These dysregulations significantly contribute to the recurrence and development of metastasis.
22,23 Existing literature has demonstrated that disrupted LPAR1 expression plays a role in promoting tumor metastasis through diverse mechanisms. For instance, phosphatidic acid (PA) has been shown to enhance cell migration by interacting with LPAR1, leading to the activation of signaling molecules such as MAPKs (ERK1-2, p38, and JNK). Additionally, it triggers the upregulation of the JAK2/STAT3 pathway.
24 In a study by Ha et al, it was revealed that LPA employs a signaling pathway mediated by Gαi2 and involves Src kinase to increase the levels of HIF1α and specific factors associated with EMT, like Slug. This process contributes to the invasive migration of ovarian cancer cells.
25 Furthermore, LPA induces EMT in ovarian tumors by activating the Wnt/β-catenin signaling pathway, which relies on the presence of β1-integrin. This novel mechanism provides a means for activating this pathway independent of mutations, contributing to the progression of epithelial ovarian cancer.
26 However, the involvement of LPAR1 in EMT and its contribution to promoting EMT in the context of PTC remain unknown. It is crucial to comprehend the role, expression pattern, and regulatory mechanisms of LPAR1 in PTC pathogenesis to establish a foundation for targeting this pathway therapeutically. This study aimed to investigate LPAR1 as a significant regulator of human thyroid cancer cell dissemination and propose that targeting LPAR1 could offer novel strategies for reducing malignancy in PTC.
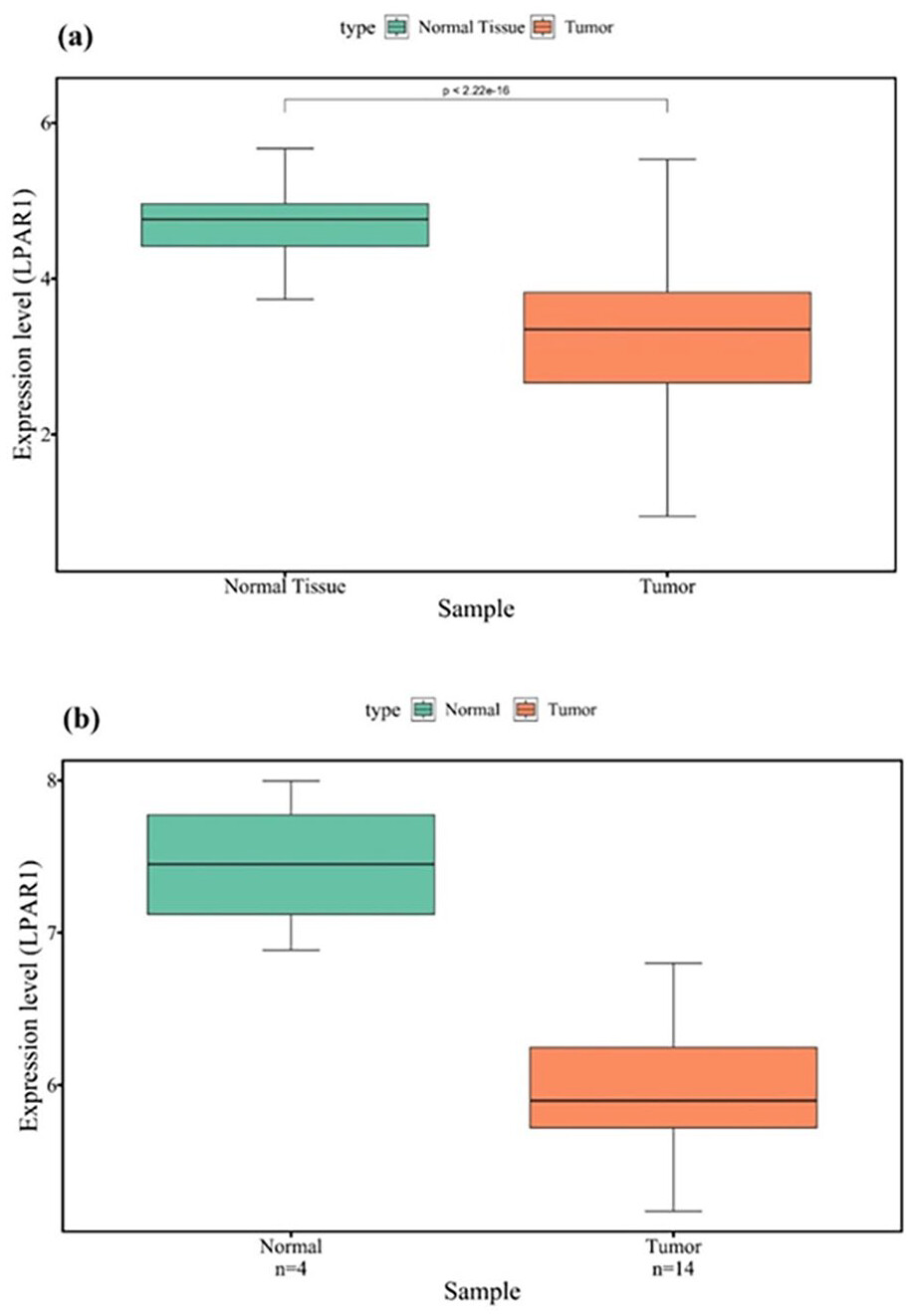
Our study aimed to identify DEGs between PTC and control cells to understand PTC pathogenesis and uncover diagnostic biomarkers and therapeutic targets, focusing on EMT. Through analysis, we identified LPAR1 and confirmed its lower expression in PTC cells. Further research revealed the involvement of the has-miR-221-5p/LPAR1 network in PTC cell EMT. The research findings on LPAR1 highlight its diverse signaling pathways and functions in different tumor cell types. Notably, LPAR1 has been reported as a negative cell motility and invasion regulator, promoting epithelial maturation.
27 In prostate cancer, LPAR1 via the downstream Gα pathways plays a role in facilitating multicellular differentiation while suppressing invasiveness. Clinical data analysis further supports that LPAR1 and Gα12/13 signaling act as metastasis suppressors, regulating the balance between cell motility, invasion, and epithelial maturation.
12LPAR1 exhibited substantial downregulation in gastric cancer tissues, displaying an inverse association with the expression of miR-501-5p. Notably, the reduction in LPAR1 expression was found to enhance both cell proliferation and migration. However, these effects were counteracted when gastric cancer cells were co-transfected with a miR-501-5p inhibitor. These findings suggest that miR-501-5p can promote cell proliferation and migration in gastric cancer by downregulating LPAR1. Therefore, the miR-501-5p/LPAR1 axis holds promise as a potential therapeutic target for the treatment of gastric cancer.
18 Conversely, our findings contrast with previous studies that indicate an elevation in LPAR1 expression in certain tumors, leading to tumor-promoting effects. For instance, it has been demonstrated that heightened levels of LPAR1 stimulate increased HIF1α levels through the Gαi2-mediated signaling pathway involving Src kinase. This, in turn, triggers EMT and facilitates invasive cell migration, accompanied by upregulated expression of N-cadherin, Slug/Snail2 in ovarian cancer cells.
25 Additionally, LPAR1 inhibition effectively prevented pulmonary metastasis of osteosarcoma cells.
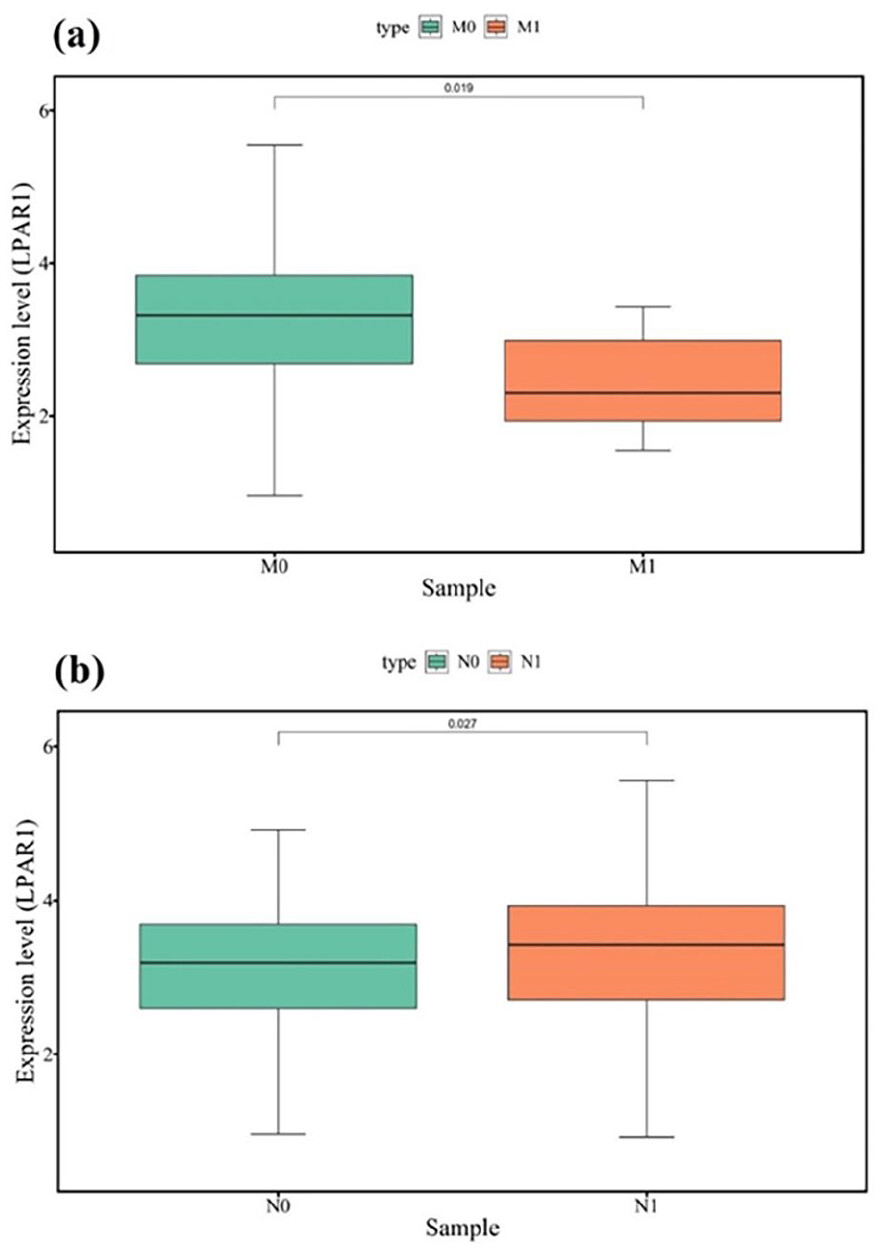
28Our bioinformatics-based study underscores the importance of LPAR1 in the EMT of thyroid cancer. We observed downregulation of LPAR1 expression in PTC tissue compared to normal tissue. Our analyses have revealed a decrease in LPAR1 expression among PTC patients with distant metastasis compared to those without metastasis. Interestingly, despite the overall reduced expression levels of LPAR1 in PTC, we observed higher expression in patients with lymph node metastasis than those without lymph node involvement. This suggests that reduced LPAR1 levels can serve as a distinguishing factor, differentiating between PTC patients with lymph node metastasis and distant metastasis and distinguishing metastatic from non-metastatic patients (
Figure 13).
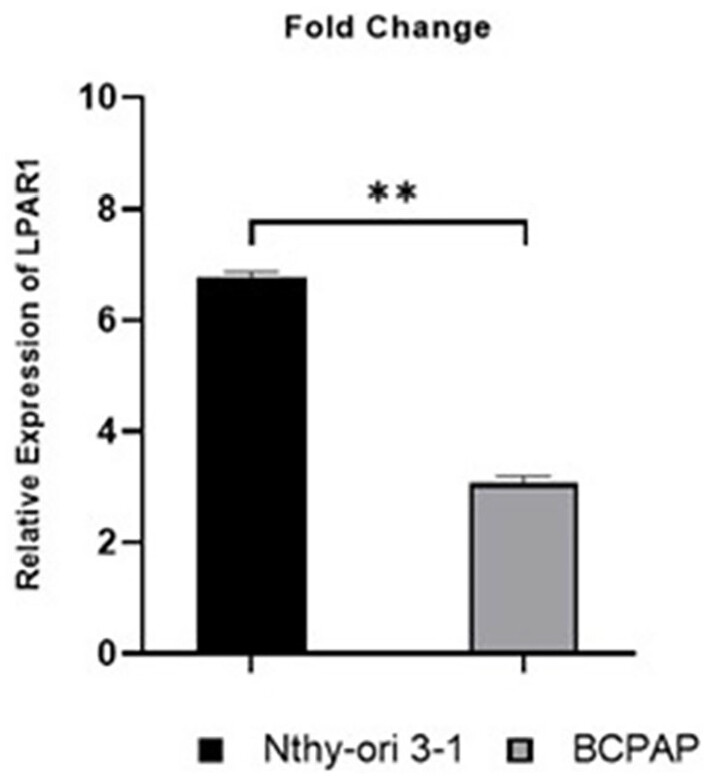
RT-qPCR experiment corroborated the findings of our in-silico analyses, demonstrating a significant downregulation of LPAR1 expression in the PTC cell line compared to the normal cell line. Additionally, significant differences in LPAR1 expression were observed between different stages of PTC, including stage I versus stage II and stage II versus stage III. Our investigation also revealed the involvement of LPAR1 in metastasis-related pathways, including phospholipase D signaling and actin cytoskeleton regulation. We found compelling evidence of LPAR1’s interaction with key proteins associated with EMT in PTC, such as ITGB4, GAB1, GNA13, ARHGEF12, PTK2, and CXCL8. The tumor microenvironment (TME) plays a pivotal role in cancer pathogenesis, which exerts its influence by modulating the balance between suppressive and cytotoxic responses in the vicinity of the tumor, thereby impacting disease outcomes.
29,30 However, the specific role of LPAR1 within the TME remains poorly understood. Using the TIMER database, we investigated the impact of LPAR1 on tumor-infiltrating immune cells in PTC. Notably, LPAR1 shows positive associations with antitumor immune cells, including effector CD4
+ T cells, macrophages, neutrophils, and dendritic cells. Conversely, there is a negative correlation between LPAR1 expression and tumor-infiltrating B lymphocytes, which is consistent with a study by Shi et al, who explored the role of LPAR1 in prostate cancer.
31,32The interaction between LPAR1 and miRNAs plays significant roles in various cancers’ pathophysiology.
33 For instance, miR-892b negatively regulates LPAR1 expression, and downregulation of miR-892b with simultaneous upregulation of LPAR1 enhances proliferation, migration, and invasion of nasopharyngeal carcinoma cells.
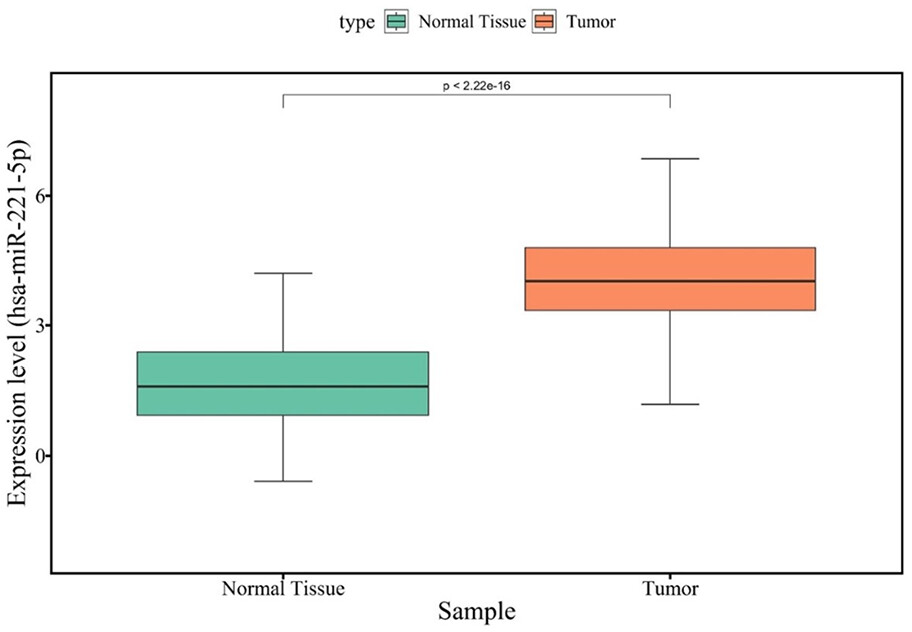
34,35 Our analysis predicts LPAR1 as a potential target of miR-221-5p, which is significantly overexpressed in PTC and inversely correlated with LPAR1 expression. The upregulation of miR-221-5p potentially suppresses LPAR1, promoting EMT in PTC. Previously, it was reported that miR-221/222 clusters are significantly upregulated in PTC compared to adjacent normal tissue. Increased levels of miR-221/222 are linked to tumor size, TNM stage, lymph node metastasis, and extra-thyroidal extension. Moreover, miR-221/222 promotes migration and invasion while inhibiting apoptosis and autophagy in PTC cells by targeting ATG10.
36 In contrast, miR-221-5p has been found to play a tumor-suppressive role in patients with prostate cancer. Loss-of-function and gain-of-function experiments have provided evidence that the overexpression of miR-221-5p reduces cell proliferation, migration, and EMT markers in vitro. Moreover, it leads to decreased tumor growth, cell extravasation, and formation of metastases in vivo.
37In summary, our study utilizing bioinformatic analyses demonstrates that LPAR1 acts as a metastasis-suppressor in PTC. Decreased expression of LPAR1 is associated with promoting EMT and metastatic behavior in PTC patients. Additionally, LPAR1 expression correlates with tumor-infiltrating immune cells, suggesting its potential as an immune-related biomarker in PTC. This highlights the potential role of LPAR1 as a prognostic biomarker for EMT and metastasis in PTC. Moreover, the findings from our study propose LPAR1 as a potential therapeutic target in PTC. As metastasis-suppressor, restoring LPAR1 expression or manipulation of its downstream signaling pathways may have therapeutic benefits in inhibiting the development of metastatic phenotype and improving patient outcomes. Hence, identifying patients with altered LPAR1 expression levels could aid in predicting their response to immunotherapeutic interventions and guiding treatment decisions. To gain a deeper understanding of the mechanisms through which LPAR1 contributes to the metastasis of PTC, further experiments conducted both in vitro and in vivo are required.